
Effect of Deastringency Treatment with CO2 on Physiological Quality of‘Hiratanenashi’ Persimmon Fruit
2021-09-27 06:47HANWeijuanCAOKunSUOYujingDIAOSongfengSUNPengLIHuaweiFUJianmin
食品科學 2021年17期
HAN Weijuan, CAO Kun, SUO Yujing, DIAO Songfeng, SUN Peng, LI Huawei, FU Jianmin,*
(1.Non-timber Forest Research and Development Center, Chinese Academy of Forestry, Zhengzhou 450003, China;2.College of Forestry, Nanjing Forestry University, Nanjing 210037, China;3.College of Forestry, Inner Mongolia Agricultural University, Hohhot 010019, China)
Abstract: In this study, the effect of CO2 deastringency treatment on the physicochemical properties of ‘Hiratanenashi’persimmon fruit was analyzed.The results revealed that after treatment with CO2, soluble solid content and fruit firmness were maintained at a high level despite a slight decrease, soluble tannin content was decreased to an acceptable level for consumption, the contents of malonodialdehyde (MDA) and polyphenols and the activity of polyphenol oxidase (PPO)were significantly reduced (P < 0.05), and total flavonoid and ascorbic acid contents and the activity of peroxidase (POD),superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX) were increased significantly (P < 0.05).This suggests that reactive oxygen species (ROS)-scavenging enzymes and non-enzymatic antioxidants can effectively scavenge the oxidative stress induced by high-CO2/hypoxic atmosphere and maintain redox balance.In addition, 9 957 and 8 984 characteristic ions were identified using metabolomics in the positive and negative ion modes, respectively, with 6 817 and 4 726 of them being annotated, respectively.In the comparison with the untreated control group, a total of 55 and 11 metabolites were found to be up- and down-regulated, respectively, including 6 down-regulated flavonoids metabolites and 14 up-regulated amino acids metabolites.Together, these results study provide valuable information on the effects of CO2-deastringency treatment on persimmon fruit and will contribute to the ongoing popularization of ‘Hiratanenashi’.
Keywords: astringent persimmon; CO2 deastringency treatment; physiological quality; metabolites; ‘Hiratanenashi’
Persimmon (Diospyros kakiThunb.) is a plant that belongs to the family Ebenaceae and has a long history of cultivation.The cultivars ofD.kakiare usually classified into four groups based on the mode of astringency loss:pollination-constant non-astringent (PCNA), pollinationvariant non-astringent (PVNA), pollination-constant astringent (PCA), and pollination-variant astringent (PVA)[1].The ‘Hiratanenashi’ persimmon is a nonaploid PCA cultivar generated from crossing reduced (n) and unreduced (2n)gametes of hexaploids with vigorous growth, a lack of seeds,enlarged cell sizes, fine flesh, and a good taste[2-3].However,the fruit are astringent at harvest because of the high soluble tannin contents in their flesh; therefore, the postharvest application of a deastringency treatment is necessary before commercialization.
The traditional methods for deastringency at harvest consist of treating fruit by over ripening, which can result in a drastic loss of firmness.Thus, fruit become very soft after astringency is removed, which seriously shortens the shelf life, and the fruit can then only be sold in local markets for a short time[4].Compared to traditional methods, CO2treatment is the most widely used deastringency technique due to its efficiency in preserving high levels of fruit firmness and user-friendliness[5-6].The effectiveness of this treatment is based on its ability toinsolubilize soluble tannins via the mediation of the acetaldehyde generated during anaerobic respiration when fruit are exposed to a high-CO2atmosphere[7-8].
Several reports revealed that the changes in physiological parameters which related to CO2treatment.Novill et al.[9]found that CO2deastringency treatment induced oxidative stress in the fruit of the ‘Rojo Brillante’ persimmon.In addition, the activities of scavenging enzymes such as superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), were increased after CO2deastringency treatment.The effects of this treatment on the firmness,flesh structure, total soluble solids content, respiration rate,ethylene production rate, and levels of vitamin C, sugars, and volatile active compounds (VOCs) have also been described for ‘Rojo Brillante’[6,10-11].Zhu Qinggang et al.[12]showed that eight transcription factors were strongly up-regulated by an artificially high CO2atmosphere in ‘Tonewase’, ‘Mopan’,and ‘Jingmianshi’ persimmon fruit.In addition, recent studies have reported that the application of a CO2-deastringency treatment may result in internal flesh browning[4,13-14], which significantly affects the economic value of fruit and greatly limits the application of CO2-deastringency technology.
The CO2-deastringency treatment has been successfully used in ‘Hiratanenashi’ fruit, with rare reports of flesh browning.Our previous studies also showed that the flesh of‘Hiratanenashi’ is not susceptible during CO2-deastringency treatment and maintains a good quality.However, few information is available regarding how physiological properties may be affected by the application of a CO2-deastringency treatment.Therefore, it is necessary to conduct a systematic study of the effects of deastringency treatment with CO2on the postharvest physiological qualities of ‘Hiratanenashi’ persimmon fruit.
The objective of this study was to determine the effects of a CO2deastringency treatment on the physicochemical properties of persimmon cv.‘Hiratanenashi’.The fruit firmness and contents of tannins (soluble and insoluble tannins), soluble solids, total flavonoids, ascorbic acid,and malondialdehyde (MDA), together with the activities of reactive oxygen species (ROS)-scavenging enzymes(peroxidase (POD), SOD, CAT, and APX) were evaluated in control and CO2-treated fruit.In addition, an untargeted metabolomic program was run to explore the effects of CO2treatment on metabolite changes in both positive electrospray ionization (ESI+) and negative (ESI-) modes.
1 Materials and Methods
1.1 Materials and reagents
‘Hiratanenashi’ persimmon fruit were harvested in Yuanyang Country, Henan Province, China at the commercial maturity stage on October 10, 2019.Forty fruit wererandomly collected from six 10-year-old healthy persimmon trees, with three biological replicates per sample.After harvest, fruit were bought back to the laboratory.
Ascorbic acid Shanghai Yuanye Bio-Technology Co.,Ltd.; Methanol and water for HPLC were of high performance liquid chromatography (HPLC) grade Sinopharm Chemical Reagent Co., Ltd.; Methanol, acetonitrile for liquid chromatography-mass spectrometry (LC-MS) were of LCMS grade Honeywell International Inc., USA; Formic acid of LC-MS grade Sigma Inc., USA.All other chemicals were of analytic grade.
1.2 Instrument and equipment
GY-4 firmness tester, TD-45 handheld refractometer Zhejiang Tuopu Co., Ltd., Ningbo; HPLC system equipped with an auto sampler (2707), a pump (1525) and UV detector(2998); Ultra-performance liquid chromatography (UPLC)Waters Co., Ltd., UK; Cary 300 UV-VIS spectrophotometer Agilent Co., Ltd., USA; TripleTOF 5600 plus High-resolution tandem mass spectrometer AB Sciex Co., Ltd., USA.
1.3 Methods
1.3.1 Material treatment
The persimmon fruits were divided into two homogeneous groups for control and CO2-deastringency treatment.For control samples (PC) were stored at room temperature without any treatment for 48 h.For treated persimmon samples (PT), the CO2treatment was conducted in sealed containers containing 99.999 9% food-grade CO2at room temperature (20-25 ℃) and 90% relative humidity for 24 h, then the fruit were stored at ambient temperature for another 24 h.Both the PC and PT samples were then peeled,and the flesh was immediately frozen in liquid nitrogen and stored at -80 ℃ for further analysis.
1.3.2 Determination of firmness and contents of soluble solids, total flavonoids, tannins, ascorbic acid, and MDA
Sample firmness was measured using a firmness tester using an 8 mm plunger before peeling.The mean of firmness values was at the average of 15 fruit and expressed as N.The soluble solids content was determined by TD-45 handheld refractometer.
The content of ascorbic acid was measured by HPLC[15].Briefly, the frozen sample was homogenized with 30 g/L metaphosphoric acid containing 8% (V/V) acetic acid.The supernatants were filtered through 0.22 μm organic filter membranes, which were then used for the HPLC analysis.Chromatography was conducted using a Syncronis C18column (250 mm × 4.6 mm, 5 μm) and a mobile phase(0.25‰,V/V, trifluoroacetic acid) of 1.0 mL/min.Detection was performed at 243 nm by a UV detector.
Total flavonoids content were measured using the AlCl3-(HAc-NaAc) method in accordance with Fan Jieping[16],and the contents of soluble and insoluble tannin were detected with Folin-Ciocalteu method in accordance with Oshida et al[17].MDA content was determined by the thiobarbituric acid method in accordance with Heath et al[18].
1.3.3 Determination of polyphenol oxidase (PPO), POD,SOD, CAT, and APX activities
For POD and PPO extraction, the frozen sample(5 g) was homogenized with 0.1 mol/L sodium acetate buffer(10 mL, pH 5.5) containing 1 mmol/L polyethylene glycol,and 4 g/100 mL polyvinyl-pyrrolidone (PVPP), 1% Triton X-100.For SOD and CAT extraction, the frozen sample (5 g)was homogenized in cold K-phosphate buffer (50 mmol/L,10 mL, pH 7.8) containing 5 mmol/LDL-dithiothreitol (DDT)and 5 g/100 mL PVPP.For APX extraction, the frozen tissue(5 g) was homogenized with 0.1 mol/L potassium phosphate buffer (10 mL, pH 7.8) containing 1 mmol/L ascorbic acid,0.1 mmol/L ethylenediamine tetraacetic acid (EDTA),and 2 g/100 mL PVPP.The supernatants of homogenates centrifuged at 12 000 r/min for 30 min at 4 ℃ were used for enzyme assays.
PPO activity was monitored according to Gibson[19]and Cao Jiankang[20]et al.with slight modifications.PPO activity was measured in a mixture of 4.0 mL sodium acetate buffer (50 mmol/L, pH 5.5) and 1.0 mL 4-methyl-catechol(50 mmol/L), 0.1 mL of the extracted enzyme was added after the mixture incubated at 25 ℃ for 5 min.Then, the absorbance at 420 nm was measured.One unit of PPO activity was defined as the amount of enzyme that caused an OD420nmincrease per minute under assay condition.
POD activity was determined according to Cao Jiankang et al.[20]with some modification.POD extract (0.5 mL) was mixed with 3 mL guaiacol solution (25 mmol/L), 0.2 mL H2O2(0.5 mol/L).The increase in absorption at 470 nm was measured and the result was expressed as U/g, where one unit of POD activity was defined as the amount of enzyme that caused an OD470nmincrease per minute under assay condition.
SOD activity measurement was assayed according to Kumar et al.[21]with some modifications.SOD extract (0.5 mL)was added to the mixture of 1.2 mL 50 mmol/L sodium phosphate buffer (pH 7.8), 0.3 mL methionine (MET)(0.13 mol/L), 0.3 mL EDTA (0.1 mmol/L), 0.3 mL nitroblue tetrazolium (NBT) (0.75 mmol/L), 0.3 mL riboflavin(0.02 mmol/L).Then the mixture was exposure to white light at 30 ℃ for 15 min, the absorbance of the reaction mixture at560 nm was measured.One unit of SOD activity was defined as the amount of enzyme that caused 50% inhibition of the reduction in NBT.
CAT activity was determined in accordance with Duan Xuewu et al.[22]with some modifications.CAT extract (0.5 mL)was added to 2.5 mL of 50 mmol/L phosphate buffer (pH 7.0)which containing 15 mmol/L H2O2.The decrease in absorbance at 240 nm were monitored, one unit of CAT activity was defined as the amount of enzyme that caused an absorbance change of 0.01 per minute under assay conditions.
APX activity was determined in accordance with Cao Jiankang et al[20].2.6 mL potassium phosphate buffer(50 mmol/L, pH 7.5, containing 0.5 mmol/L ascorbic acid and 0.1 mmol/L EDTA) was added into 0.1 mL of enzyme extract, then the reaction was started by adding 0.3 mL H2O2solution (2 mmol/L).The decrease in absorbance at 290 nm were monitored, one unit of APX activity was defined as the amount of enzyme that caused an absorbance change of 0.01 per minute under assay conditions.
1.3.4 Metabolite extraction and untargeted LC-MS analysis conditions
A portion (100 mg) of each collected sample was extracted by 120 μL of precooled 50% (V/V) methanol,vortexed for 1 min, and incubated at room temperature for 10 min.The extraction mixture was then stored overnight at -20 ℃.After centrifugation at 4 000 ×gfor 20 min, the supernatants were transferred into new 96-well plates and stored at -80 ℃ prior to LC-MS analysis.In addition, pooled quality control (QC) samples were prepared by combining 10 μL of each extraction mixture.
A 4-μL sample was injected into an ultra-performance liquid chromatography (UPLC) system equipped with an ACQUITY UPLC T3 column (100 mm × 2.1 mm, 1.8 μm) for reverse-phase separation.The column oven was maintained at 35 ℃.The flow rate was 0.4 mL/min, and the mobile phase consisted of solvent A (water, 0.1% formic acid)and solvent B (acetonitrile, 0.1% formic acid).The elution gradient was set as follows: 0-0.5 min, 5% B; 0.5-7 min, 5%to 100% B; 7-8 min, 100% B; 8-8.1 min, 100% to 5% B; and 8.1-10 min, 5% B.
A high-resolution tandem mass spectrometer was used to detect metabolites eluted from the column in both ESI+and ESI-modes, each ion was identified by combining the retention time andm/zdata.The QC sample was used to evaluate the stability of the LC-MS during the whole acquisition process.
1.4 Statistical analysis
The experiments were performed with an absolutely randomized design, and the results were expressed on a fresh weight basis.For the untargeted metabolomics analysis, Studentt-tests were conducted to detect differences in metabolite concentrations between two groups.ThePvalue was adjusted for multiple tests by measuring the false discovery rate using the Benjamini-Hochberg test.For the fruit firmness, components and enzymes, all the data were analyzed with SPSS 23.0 software.Two-way analysis of variance was used to assess the data, and Duncan’s multiple comparisons test was used to test for significant differences.Differences atP< 0.05 were defined as significant.
2 Results and Analysis
2.1 Effects of CO2-deastringency treatment on physiological qualities of ‘Hiratanenashi’ persimmon fruit
Fruit firmness is an economically significant property that determines the quality of persimmons when they are marketed after removing of astringency.The firmness of the control and astringency-removed persimmon fruit is presented in Fig.1A.The firmness of treated samples significantly decreased compared to control samples, with the result suggesting that fruit firmness was affected by the treatment of CO2-deastringency.However, ‘Hiratanenashi’fruit still maintained high levels of firmness ((68.92 ± 2.29) N)with a high commodity value after the astringency was removed by CO2treatment.
Insoluble and soluble tannin levels are shown in Fig.1B and Fig.1C, respectively.The soluble tannin content in treated fruit was (1.15 ± 0.02) g/kg, the threshold value of astringency was 0.2%[23], which indicated that the CO2-deastringency treatment used in this study was effective.The soluble tannin content decreased from (7.10 ± 0.04) g/kg in the control to (1.15 ± 0.02) g/kg in treated samples, whereas the insoluble tannin content increased from (6.57 ± 0.20) g/kg in the control to (13.55 ± 0.32) g/kg in the treated samples.These changes may be that the soluble tannins accumulated and converted into insoluble forms, with the subsequent impact of acetaldehyde being generated during the CO2-deastringency treatment, as previously reported[8,24].
As shown in Fig.1D, the content of soluble solids decreased from (21.3 ± 0.46)% in the control to (14.7 ± 0.75)%in treated samples.Soluble tannins are included in the soluble solids; thus, the decrease in soluble solids content after It has been reported that CO2-deastringency treatment can affect oxidative stress on fruit, and ROS levels can increase rapidly after CO2treatment[25].The effective destruction of ROS requires several ROS-scavenging enzymes, such as SOD, POD, CAT, and APX[9], to act concomitantly with non-enzymatic antioxidants, such as ascorbic acid and flavonoids[25].As shown in Fig.1E-H, the SOD, POD, CAT,and APX activities in treated samples increased significantly compared with those in the control.SOD activity in treated sampleis 2.01 times of control, whereas POD activity increased by approximately 1.28 times.CAT activity in astringency-removed persimmon fruit was (22.30 ± 1.02) U/g,which was about 1.90 times of control.APX activity in astringency-removed fruit was (135.17 ± 3.76) U/g, which was much higher than control.As shown in Fig.1I, J, levels of total flavonoids content and ascorbic acid content were both significantly increased compared with the control.deastringency treatment may be mainly due to the reduction of soluble tannin content[4].
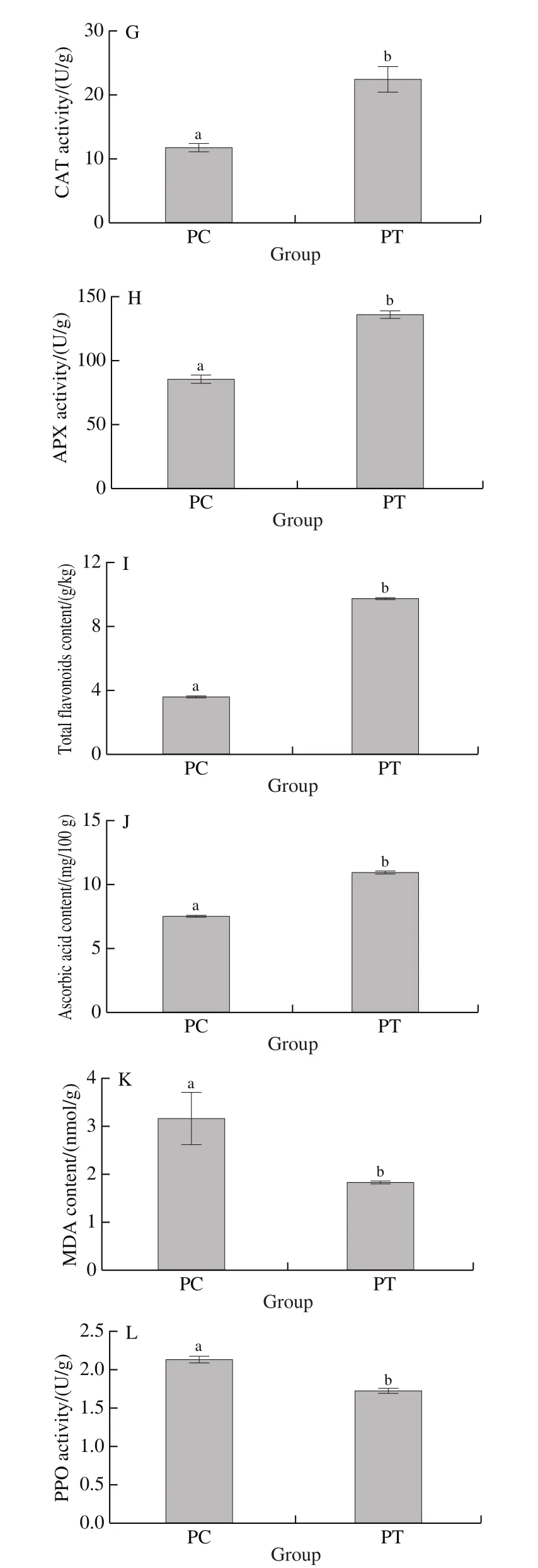
Fig.1 Physiological parameters of control (PC) and CO2-treated (PT)persimmon fruits
As shown in Fig.1K, L, The MDA content decreased from (3.14 ± 0.56) nmol/g in the control to (1.82 ± 0.06) nmol/g in treated samples; PPO activity in the control was(2.12 ± 0.06) U/g, which was about 1.24 times of the CO2treated sample.MDA content and PPO activity were both significantly decreased compared with the control.
2.2 Results of metabolomic analysis and identification of altered metabolites
To determine the changes in metabolites after CO2treatment, an untargeted metabolomic program was operated in both positive (Pos) and negative (Neg) ionization modes.As shown in Fig.2, total ion chromatograms were generated for all samples in order to examine the quality of MS data,which revealed that there was a high level of overlap,indicating that the sample preparation and instrument state met the required standards; The peaks of the detected substances not only overlapped but also differed, so that different metabolites were identified as being obtained from control and treated samples.
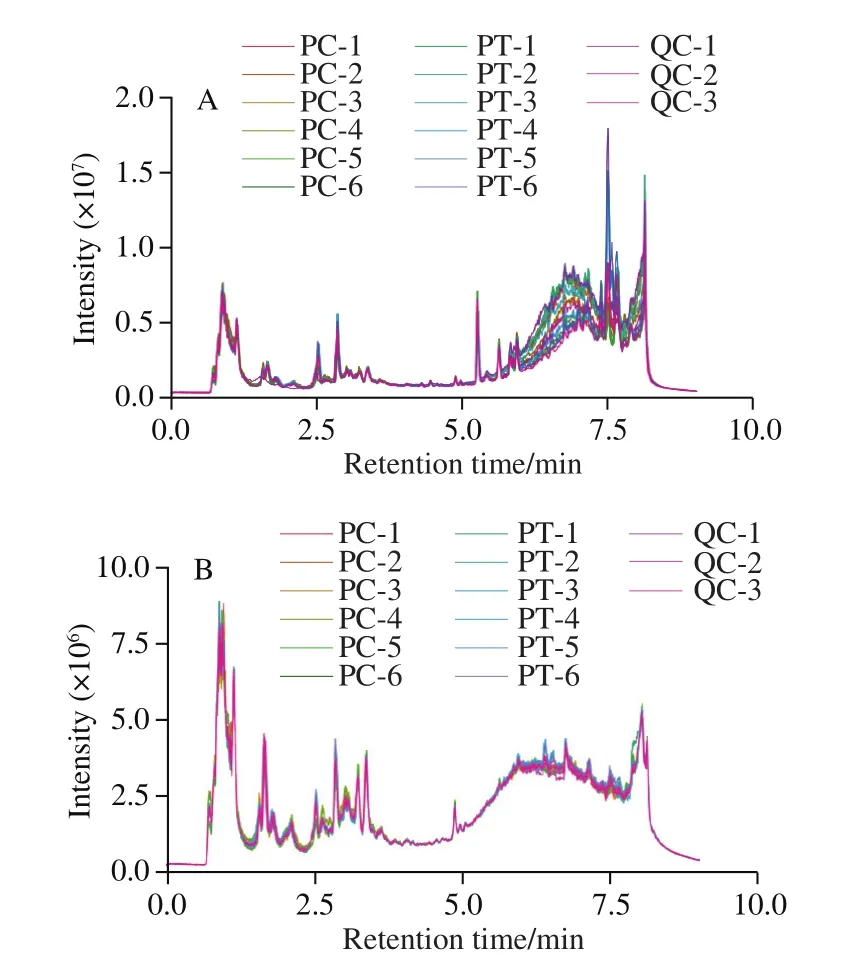
Fig.2 Total ion current chromatograms of metabolites in control and CO2-treated persimmon fruits in the positive (A) and negative (B) ion modes
A total of 9 957 and 8 984 ion features were obtained using the Pos and Neg modes, respectively; Of which 6 817 and 4 726, respectively, were annotated.832 metabolites were identified using an in-house fragment spectrum library of metabolites (Table 1).All of the 517 and 315 metabolites obtained from the MS2identification were assigned using the human metabolome database (HMDB) (HMDB annotation shown in Fig.3).Among these, “lipids and lipid-like molecules” (Pos, 175; Neg, 124) and “Organic acids and derivatives” (Pos, 53; Neg, 34) were the two largest groups.According to the Kyoto encyclopedia of genes and genomes(KEGG) classification map, the top 20 KEGG pathways were identified and the five largest number of metabolites was related to “Metabolic pathways”, “Biosynthesis of secondary metabolites”, “Biosynthesis of plant secondary metabolites”,“ABC transporters”, “Biosynthesis of amino acids” in KEGG (Fig.4).
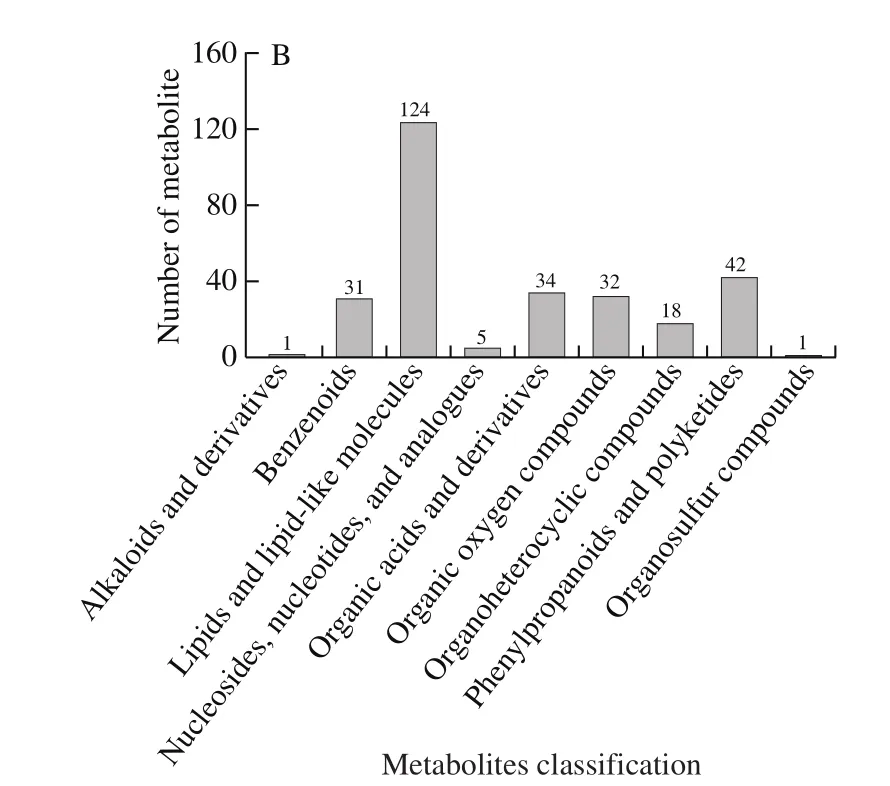
Fig.3 Classification of metabolites obtained from HMDB
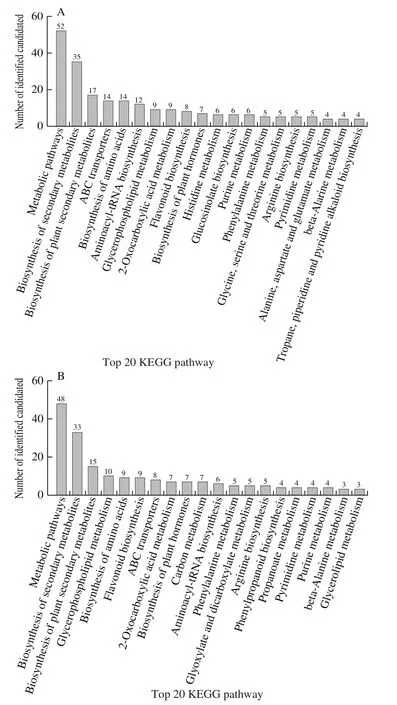
Fig.4 Top 20 KEGG pathway classifications
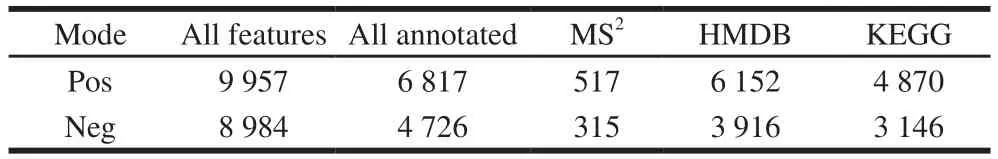
Table 1 Statistical results of metabolite annotation
The metabolic profiles of the control and treated persimmon fruit were analyzed using partial least-squares discriminant analysis (PLS-DA) in both the ESI+and ESI-modes.The PLS-DA scores plot obviously differentiated among the two groups as shown in the Fig.5.There were no significant outliers in the PLS-DA scores plot, suggesting that the samples were of high quality.The values ofR2andQ2were, respectively, 0.999 5 and 0.929 7 for the ESI+mode and 0.999 8 and 0.960 9 for ESI-mode.These data indicated that the effectiveness of the models was satisfactory, and the data obtained were reliable according to these models.
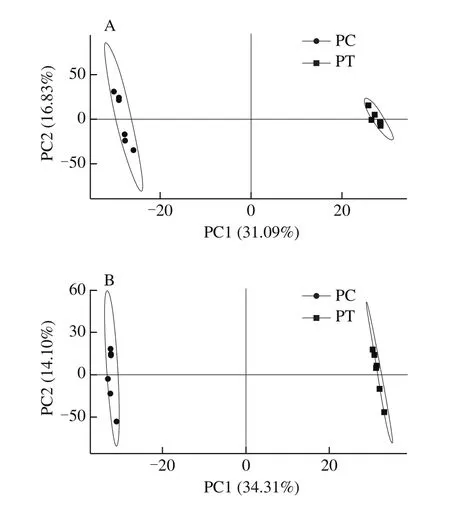
Fig.5 Partial least-squares discriminant analysis (PLS-DA) score plots for metabolite profiles of control (PC) and CO2-treated (PT) persimmon fruits in the positive (A) and negative (B) ion modes
The metabolites that differed between the control and CO2-treated groups were compared based on the fold change,VIP value andPvalue.A total of 55 and 11 metabolites were respectively found to be up- and down-regulated in the control and CO2-treated samples (Table 2).In addition,six metabolites that differed significant between groups(catechin, epicatechin, delphinidin, kaempferol, taxifolin,and dihydrorobinetin) were identified as flavonoids among the MS2metabolites.A total of 14 amino acids were identified: leucine,L-norleucine, citrulline, phenylalanine,L-tyrosine,L-histidine, ornithine, glutamine,L-5-oxoproline,tryptophan,L-tryptophan,L-(-)-phenylalanine, (±)-erythroisoleucine, and glutamic acid (Table 2).Lysophospholipids are recognized as membrane phospholipid metabolites.As shown in Table 2, LysoPC, LysoPE, LysoPS, and LysoPA contents increased significantly, whereas LysoPG content decreased after CO2treatment.Additionally, 2-phenylacetamide, 2-hydroxycinnamic acid, and 3-hydroxycinnamic acid, which are involved in phenylalanine metabolism, were up-regulated.
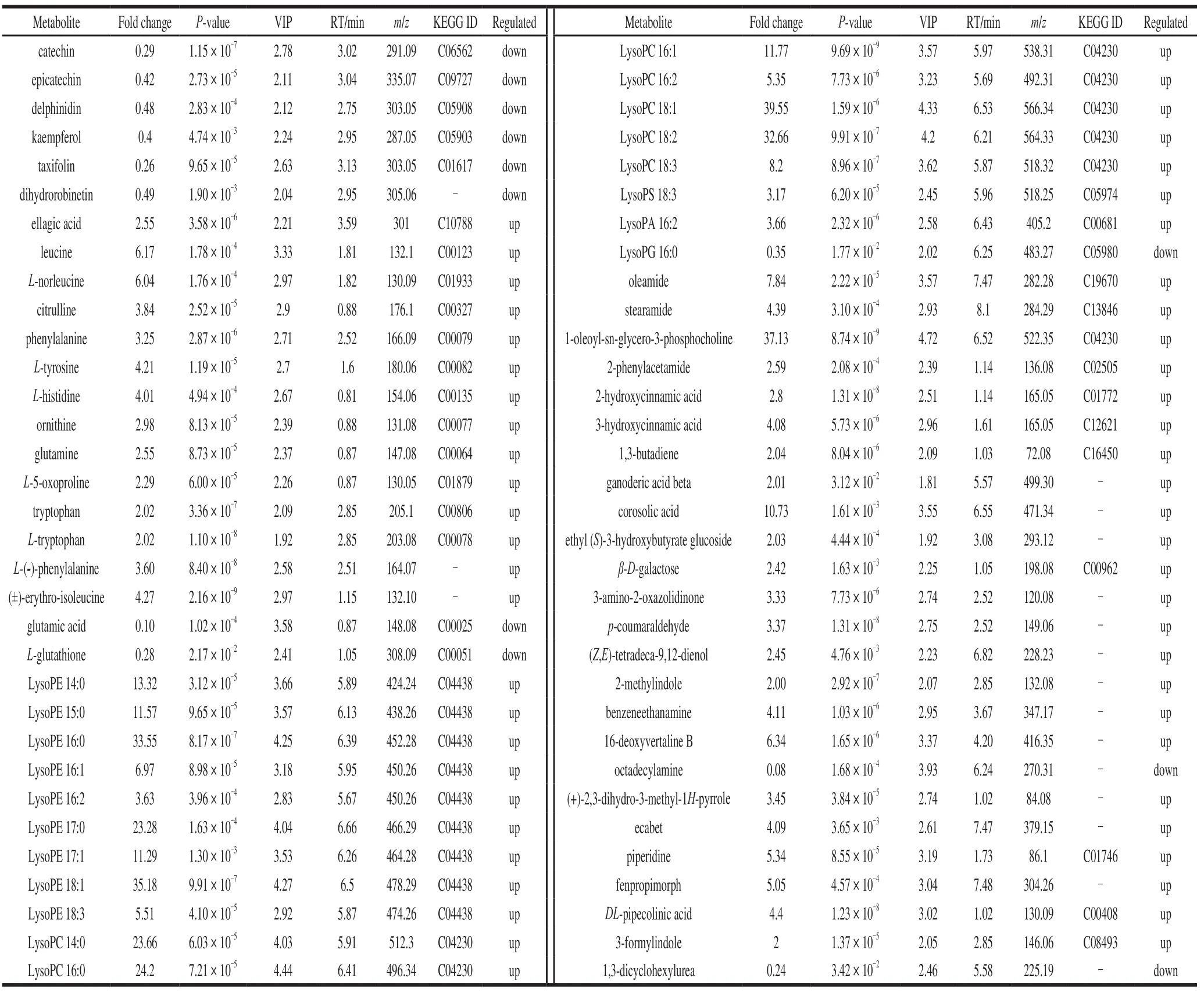
Table 2 Comparison of metabolites between control (PC) and CO2-treated (PT) fruits
3 Discussion
The effectiveness of a CO2-deastringency treatment relies on exposing fruit to anaerobic conditions.We found that the CO2-deastringency treatment used in this study was effective because the soluble tannin content was (1.15 ± 0.02) g/kg under the threshold value of 0.2%.The soluble tannin content decreased after CO2deastringency treatment, whereas insoluble tannin content increased.These changes may be due to the soluble tannins being polymerized to insoluble tannins,with the acetaldehyde generated and accumulated during the CO2deastringency treatment, as previously reported[8,24].Decrease in firmness have often been linked to a reduction in the soluble tannins content which are responsible for the astringency[6,26]; However, the result of present study showed that CO2-deastringency treatment preserved high levels of fruit firmness.
The soluble solids content decreased from (23.30 ± 0.46)%to (14.70 ± 0.75)% after the CO2-deastringency treatment.The loss of astringency after treated by CO2treatment due to the conversion of soluble tannins into insoluble form, the soluble tannins are included in the measurement of soluble solids.Therefore, it was concluded that the decrease of soluble solids content was related to the loss of astringency following CO2treatment.
Persimmon fruit treated with high concentrations of CO2undergo hypoxic stress that induces the accumulation of ROS[25].The SOD, POD, APX, and CAT are capable of scavenging ROS in plants[27].SOD provided the first enzymatic line of defense against ROS through catalyzes the dismutation of O2-·to non-toxic O2and less toxic H2O2,H2O2can be removed by POD, CAT, and APX.As a result of membrane lipid peroxidation, MDA content is used as an important indicator to reflect membrane damage and as a suitable biomarker for oxidative stress[27].In most higher plants, PPO plays a central role in fruit browning[28].In this study, SOD, POD, CAT, and APX activities and contents of ascorbic acid and flavonoids increased significantly after CO2-deastringency treatment.The MDA content and PPO activity decreased after treatment with CO2, with the results suggesting that ROS-scavenging enzymes and non-enzymatic antioxidants can effectively maintain the balance of redox reactions and protect cell membranes and intracellular proteins from oxidative damage inside the fruit during the CO2-deastringency treatment.
In plant metabolomics, the roles of metabolite changes in various tissues were examined[29].Because metabolites are the final products of the cell biological regulation process,changes in metabolites and their levels are regarded as the ultimate response of plants to environmental stresses[30].In the comparison between control and treated samples, a total of 55 and 11 metabolites were found to be up- and downregulated, respectively.
Catechin, epicatechin, delphinidin, kaempferol, taxifolin,and dihydrorobinetin were down-regulated, whereas ellagic acid, which is the dimer derivative of gallic acid, was upregulated in the treated group.These changes may be due to the soluble tannins being accumulated and converted to insoluble tannins after the CO2-deastringency treatment.
Amino acids are considered to be precursors and constituents of proteins.Many amino acids also play significant roles in plant growth and the ascorbic-acid response to various stresses, including acting as regulatory and signaling molecules[31-32], and several studies have demonstrated that high levels of amino acids accumulate under abiotic stress[33-36].In this study, the levels ofL-norleucine, citrulline,L-phenylalanine,L-tyrosine,L-histidine, ornithine, glutamine,L-5-oxoproline, tryptophan,L-tryptophan,L-(-)-phenylalanine, and (±)-erythro-isoleucine were up-regulated under a high-CO2/hypoxic atmosphere.One possible reason for this is that high CO2concentrations could induce the synthesis of amino acids and the degradation of proteins in persimmon fruit.Glutamate not only occupies a central position in amino-acid metabolism but also affects or regulates some important physiological processes in plants[37-38].Glutamate was down-regulated in this study,suggesting that it may serve as a substrate or precursor in amino-acid metabolism.L-glutathione is an important component of the antioxidant network, which consists of lowmolecular weight antioxidants that balance redox reactions[39].The mechanism of its down-regulation in this study needs to be confirmed in the next stage of the research.
4 Conclusion
To summarize, the CO2-deastringency treatment affected the physicochemical properties of persimmon cv.‘Hiratanenashi’.The astringency was removed via the reduction of soluble tannins in parallel with the accumulation of insoluble tannins.SOD, POD, CAT, and APX activities and levels of total flavonoids and ascorbic acid increased significantly, whereas MDA content and PPO activity decreased, indicating that the ROS-scavenging enzymes and non-enzymatic antioxidants can effectively maintain the balance of redox reactions under high-CO2/hypoxic conditions.In addition, the metabolites in the treated and control groups were compared using an untargeted metabolomic program.A total of 55 and 11 metabolites were found to be up- and down-regulated, respectively.In total,6 flavonoids and 14 amino acids metabolites were found to be involved in deastringency and low-oxygen stress.These results provide valuable information for future studies on the effects of CO2-deastringency treatment on ‘Hiratanenashi’persimmon fruit.
